
FOTOSSÍNTESE
SIMPLIFICADA E RESUMIDA
Maximiliano
Mendes
Podemos classificar os organismos em dois grupos, quanto à forma como obtêm alimentos:
Heterótrofos: não são capazes de produzir as substâncias orgânicas
que lhes servem de alimento a partir de compostos inorgânicos simples. Logo,
têm de ingeri-las. É o caso, por exemplo, dos animais. Lembre-se de que as substâncias
orgânicas são aquelas constituídas primordialmente de CHONSP: carbono,
hidrogênio, oxigênio, nitrogênio, enxofre e fósforo. (*O CO2 não
é considerado uma substância orgânica).
Autótrofos: são capazes de produzir as substâncias orgânicas que
lhes servem de alimento. Bons exemplos de organismos autótrofos são as plantas,
as algas verdes e as cianobactérias, todos fotossintetizantes. Há também os
procariontes quimiossintetizantes.
Podemos considerar que todos os ecossistemas necessitam da presença de organismos autótrofos, que atuarão como os produtores (1º nível trófico). No caso, produtores de substâncias orgânicas para eles e para quem os consome, os consumidores (2º, 3º... níveis tróficos), heterótrofos.


A fotossíntese é um mecanismo de síntese de substâncias orgânicas a partir
de CO2 e energia luminosa. De certa forma, podemos imaginar a
fotossíntese como um processo que se dá no sentido contrário ao da respiração
celular, como se pode ver nas equações simplificadas abaixo:
Respiração celular: Glicose (“alimento”) + 6 O2 --> 6 CO2 + 6 H2O + Energia potencial química.
Fotossíntese: 6 CO2 + 6 H2O + Energia luminosa --> Glicose (“alimento”) + 6 O2.
Respiração celular: Glicose (“alimento”) + 6 O2 --> 6 CO2 + 6 H2O + Energia potencial química.
Fotossíntese: 6 CO2 + 6 H2O + Energia luminosa --> Glicose (“alimento”) + 6 O2.
Veja que os produtos de um processo podem ser considerados como os
reagentes do outro.

Fase "clara" ou
etapa fotoquímica:
Essa etapa ocorre nas membranas tilacoides e pode ser chamada de fase "clara" por depender diretamente da energia luminosa, ou seja: só ocorre na presença de luz. O processo utiliza pigmentos, moléculas capazes de absorver a luz visível. Na fotossíntese, os principais tipos de pigmentos envolvidos são as clorofilas, localizadas em dois tipos de complexos proteicos, os chamados fotossistemas II e I (PS II e PS I).
Inicialmente, as moléculas de clorofila dos fotossistemas II absorverão a
energia luminosa e isso fará com que os seus elétrons sejam excitados. Elétrons
excitados se tornam instáveis e serão transferidos com alta energia para bombas
de íons H+, também localizadas nas membranas tilacoides.
A luz pode se comportar como radiação eletromagnética, propaga-se por ondas eletromagnéticas, e também como
partículas chamadas fótons de luz, pequenos pacotes de energia. Imagine que os fótons sejam como bolas de
sinuca muito pequenas e os elétrons também. Em um jogo de sinuca, quando uma
bola em movimento atinge uma bola parada, transfere energia para ela e faz com
que ela se mova. No nível subatômico, quando os fótons de luz atingem os
elétrons das clorofilas, transferem energia para eles, o que os excita e permite
que sejam enviados com alta energia, como bolinhas de sinuca, para outras
moléculas que os captam, como exemplos, veremos coenzimas captadoras de
elétrons de alta energia, bombas de íons H+ e outras máquinas
proteicas.

Os elétrons perdidos pelas clorofilas dos fotossistemas II serão
repostos pela H2O, em um processo chamado de fotólise da água (veja
a figura):
H2O --> 2 H+ + ½ O2 +
2 e-
Atenção para o fato de que o O2 gerado pela fotossíntese é proveniente da H2O e não do CO2!
Como se pode ver na figura acima, da cadeia de transporte de elétrons da
etapa fotoquímica, a luz incide sobre as clorofilas de ambos os fotossistemas e
em ambos os casos ocorre a liberação de elétrons de alta energia.
Os elétrons que transferiram energia para as bombas de H+ serão transferidos para as moléculas de clorofila dos fotossistemas I a fim de repor os elétrons que elas estão perdendo sendo enviados para a próxima fase da fotossíntese.
Enfim, é uma cadeia de transporte de elétrons na qual se pode imaginar
que inicialmente os elétrons partem da água.
Vimos que ocorre o bombeamento de íons H+ e que isso
gera uma diferença de concentração entre o interior
dos tilacoides (concentração maior) e o estroma do cloroplasto
(concentração menor). Assim, os íons H+ tendem a atravessar as
membranas tilacóides pela ATP sintase, basicamente a mesma da respiração
celular: uma permease com uma turbina acoplada que pode girar e fosforilar ADP
para formar ATP, graças à energia fornecida pela passagem dos íons H+ através
dela:

A quantidade de ATPs gerados depende da intensidade luminosa,
disponibilidade de água, temperatura e outros fatores, por isso não é
necessário decorar as quantidades geradas.
É importante perceber que a fase clara da fotossíntese é similar ao que
ocorre na cadeia de transporte de elétrons e fosforilação oxidativa
mitocondriais (aqui, chamada fotofosforilação), só que ao invés de serem as
etapas finais, na fotossíntese, essas são as etapas iniciais. Note também que
na fotossíntese se gera O2 (a partir da quebra água) e na
respiração se consome O2.
A etapa fotoquímica produz ATP e elétrons de alta energia para
a próxima fase (fase "escura" ou de fixação do carbono).
Esses elétrons de alta energia em trânsito participam de reações de
oxidação-redução e são transferidos de uma espécie química para outra. Então não
vagam livres pelas soluções celulares, eles são captados por substâncias
aceptoras de elétrons, que, ao captá-los são reduzidas e ao transferi-los, são
oxidadas. Ao término da fase fotoquímica, os elétrons de alta energia a serem
utilizados na próxima fase da fotossíntese são captados e transportados por uma
coenzima chamada dinucleotídeo de adenina e nicotinamida fosfato (NADP+)
conforme a reação abaixo:
NADP+ + H+ + 2 e-
--> NADPH
O NADP+ é a forma oxidada e o NADPH é a forma reduzida
(captou os elétrons). O NADPH, posteriormente sofre oxidação e transfere os
elétrons de alta energia previamente captados. Veja também que o NADP+
também capta H+, para balancear as cargas negativas dos elétrons.
A imagem abaixo apresenta a mesma cadeia de transporte de elétrons da
fotossíntese, porém, de forma mais detalhada, mostrando várias das substâncias
que atuam nas reações de oxidação-redução:

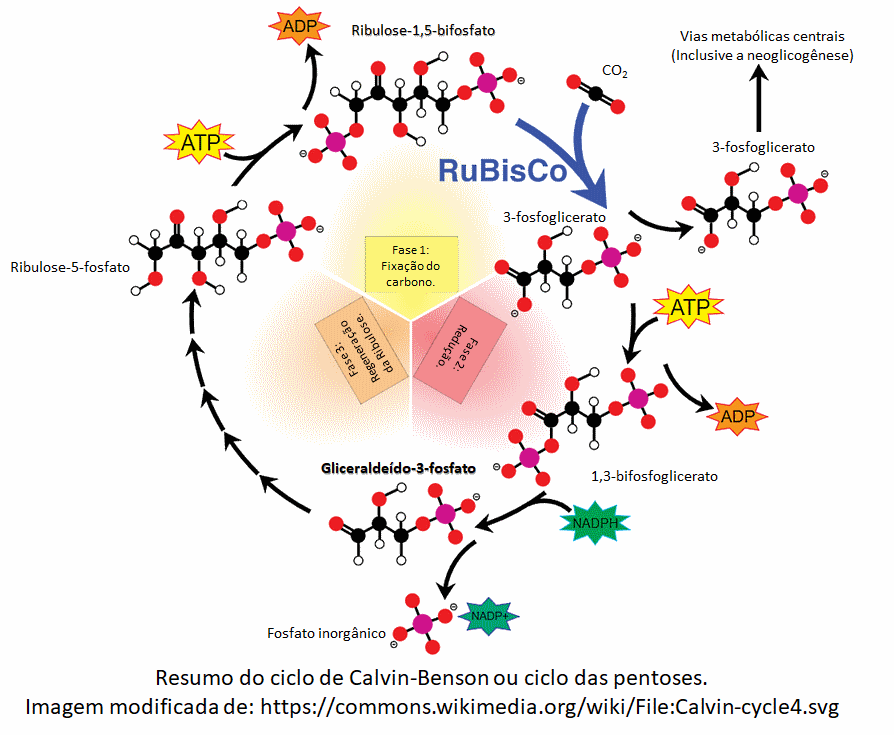
Fase "escura" ou de fixação do
carbono:
Também chamada de fase puramente química. Consiste em um ciclo de
reações, o ciclo de Calvin-Benson (ou ciclo das pentoses), que ocorre
no estroma dos cloroplastos e não depende diretamente da luz, mas depende
diretamente dos produtos da fase clara: ATP e os elétrons de alta energia
transportados pelo NADPH que os havia captado. Logo, por não depender
diretamente da luz, o ciclo de Calvin pode ocorrer na presença e na
ausência de luz (de dia e de noite). Lembrando que, na ausência de luz, o ciclo
persiste enquanto durarem os produtos da fase clara, então, as plantas que não
recebem iluminação adequada podem morrer.
Basicamente, o ciclo de Calvin utiliza moléculas de CO2 para
a geração de uma determinada substância orgânica, com o uso de ATPs e elétrons de
alta energia (equação simplificada):
6 CO2 + 18 ATP + elétrons de alta energia --> 2
gliceraldeído-3-fosfato (C3H7O6P).
Uma forma de escrever a equação simplificada com os NADPH pode ser:
3 CO2 + 9 ATP + 6 NADPH -> C3H6O3-fosfato + 9 ADP + 8 Pi + 6 NADP+
As moléculas de gliceradeído-3-fosfato ou fosfogliceraldeído, abreviadas
como PGAL ou G3P, podem ser utilizadas pelas células para gerar outras
substâncias que lhes servem de alimento e também para os organismos que as consomem,
como os herbívoros. Como exemplo, um processo chamado neoglicogênese utiliza
dois PGAL para gerar uma molécula de glicose.
O processo tem o nome de ciclo, pois um dos reagentes iniciais é também
um dos produtos finais, no caso, uma molécula chamada de ribulose-1,5-bifosfato.
A imagem abaixo resume o processo:
Veja também:
REFERÊNCIAS
Amabis & Martho. Biologia das Células. Moderna. 2010.
Campbell,
Reece et al. Biologia. 8ª Ed. Artmed.
Catani et al. Ser Protagonista – Biologia – Vol. 1. Edições
SM. 2009.
Polkinghorne,
J. Quantum Physics and Theology – An Unexpected Kinship. Yale
University Press. 2007.
Na internet:
https://www.youtube.com/watch?v=eo5XndJaz-Y
https://www.youtube.com/watch?v=0UzMaoaXKaM
(Esses dois últimos vídeos, TED-Ed, têm legendas em língua portuguesa disponível).













{kind=link}