
DESENVOLVIMENTO EMBRIONÁRIO DOS ANIMAIS
Maximiliano Mendes
1. O início da vida de um animal
A fertilização é o processo no qual ocorre o encontro e a união entre um gameta
feminino e um masculino. A vida de um novo indivíduo da espécie tem início após a fertilização com a formação do zigoto: a célula que contêm material genético dos dois
progenitores.

O zigoto irá se dividir por mitose, originando células chamadas blastômeros e essas, por sua vez,
continuarão a se dividir e originarão outros tipos celulares via diferenciação celular.
Antes de iniciarmos o estudo
do desenvolvimento de um animal, veremos alguns conceitos importantes.
2. Vitelo
Tanto o zigoto quanto o
gameta feminino podem ser chamados de célula
ovo (ou simplesmente ovo). Isso porque o gameta feminino, o óvulo, é uma
célula muito maior que um espermatozoide, logo, o zigoto tem praticamente o
mesmo tamanho de um óvulo. Lembrando que a célula na qual o espermatozoide penetra é chamada ovócito II. Essa célula termina a meiose II e gera um óvulo, com o espermatozoide dentro, e um corpúsculo polar, após a fertilização.
Os ovos armazenam substâncias
nutritivas (proteínas e lipídios) no citoplasma: o vitelo. Esse vitelo tem como função nutrir as células do embrião em
desenvolvimento até que ele consiga obter alimento por conta própria. Um
detalhe interessante sobre o vitelo é que, quanto maior for a quantidade de
vitelo em determinada região do citoplasma, mais difícil é dividir essa região,
você pode tentar imaginar o vitelo como se fosse manteiga congelada: não é muito fácil cortar com uma faca de plástico.
2.1. Tipos de células ovos em relação à quantidade de
vitelo
Isolécitos ou microlécitos:
têm pouco vitelo distribuído de forma mais ou menos homogênea pelo citoplasma,
alguns inclusive são chamados de alécitos, ou seja, sem vitelo. Presente nos
ovos de mamíferos placentários, equinodermos, moluscos, anelídeos, nematódeos e
platelmintos.
Heterolécitos: possuem
uma quantidade relativamente grande de vitelo, distribuída de forma desigual no
citoplasma do ovo. Uma região apresenta pouco vitelo (o polo animal) e outra
apresenta bastante vitelo (o polo vegetativo). Presente em anfíbios, nas lampreias e os peixes pulmonados.
Telolécitos ou Megalécitos: ovos grandes, com bastante vitelo, que ocupa
praticamente todo o citoplasma, exceto uma região restrita a um pequeno disco
superficial. Presente em répteis, aves, alguns peixes, moluscos cefalópodes
(polvos, lulas e sépias) e mamíferos prototérios (os monotremados, como o ortnitorrinco).
Centrolécitos:
grande quantidade de vitelo localizado na região central da célula (onde também
se encontra o núcleo). O citoplasma é restrito a uma pequena região logo abaixo
da membrana plasmática. Presente
mais comumente em artrópodes.

2.2. Tipos de segmentação ou clivagem:
Segmentações ou clivagens são
os nomes dados às divisões celulares (mitoses) pelas quais as células
embrionárias passam. Podem ser classificadas de quatro formas:
Holoblástica ou total: a célula ovo pode ser completamente dividida de um polo
ao outro. Isso se deve ao fato de que no citoplasma não há vitelo em grandes
quantidades.
Meroblástica ou parcial: devido à grande quantidade de vitelo presente no
citoplasma, somente parte dele sofre clivagem.
Igual: os blastômeros originados a partir das clivagens têm tamanhos praticamente iguais. Isso pode ser devido à distribuição uniforme do vitelo no citoplasma da célula ovo.
Desigual: os
blastômeros originados a partir das clivagens têm tamanhos distintos
(micrômeros e macrômeros), o que pode ser devido à distribuição não uniforme do
vitelo pelo citoplasma da célula ovo.

3. O desenvolvimento embrionário dos animais:
Estudaremos o processo de
desenvolvimento embrionário dos animais tendo como modelo um protocordado
marinho chamado anfioxo (Branchiostoma
lanceolatum). Esse animal é utilizado como modelo e exemplo, pois os processos do desenvolvimento são basicamente os mesmos da nossa espécie, têm menos detalhes e, portanto, podem ser
entendidos com menos dificuldades.

3.1. Estágios do desenvolvimento dos animais:
A figura abaixo mostra quais
são os estágios do desenvolvimento de um animal, partindo do zigoto e, lembrando, usando o anfioxo como modelo. Detalhe: o
zigoto não é um embrião, o animal passa para o estágio embrionário após o início das clivagens, as primeiras mitoses.
3.2. Segmentação ou clivagens: Zigoto > Mórula > Blástula.
Processo no qual um zigoto se
desenvolve em mórula e depois em blástula. Há aumento do número de células,
graças às divisões, porém, não há aumento do volume do embrião. Isso se deve ao
fato de que as divisões celulares ocorrem muito rapidamente, então as células nem têm tempo para crescer.

As células originadas são
chamadas de blastômeros, sendo que
podem ser de dois tipos:
Micrômeros:
menores e originadas de regiões com menores concentrações de vitelo.
Macrômeros:
maiores e originadas de regiões com maiores concentrações de vitelo.
Após algumas divisões o embrião se desenvolve em uma Mórula: estrutura
maciça constituída de um amontoado de células (blastômeros) similar a
uma amora. As células ainda não estão diferenciadas.
Prosseguindo as divisões (clivagens) o embrião se desenvolve em uma Blástula: uma esfera cujo interior é preenchido por liquido. O interior preenchido por líquido é chamado blastocela (a cavidade da blástula) e a superfície é constituída por uma camada de células chamada blastoderme. As células ainda não estão diferenciadas. O líquido não tem nome, chama-se líquido mesmo.

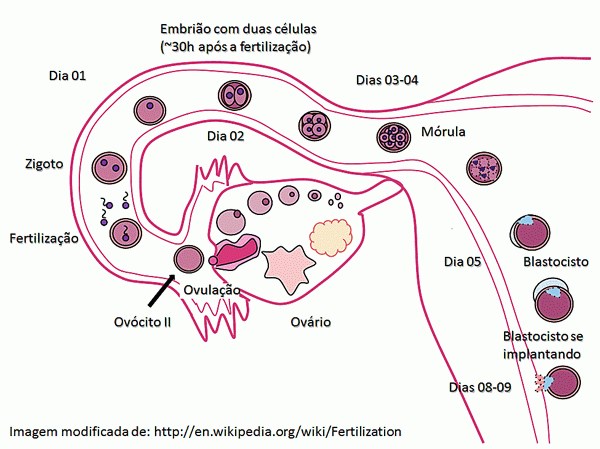
A blástula dos mamíferos é
chamada de blastocisto. O embrião
humano se encontra nesse estágio de desenvolvimento aproximadamente cinco dias
após a fertilização. Ainda nesse estágio, de sete a nove dias após a
fertilização, o embrião se implanta no endométrio do útero. Pode se retirar
células tronco embrionárias para serem utilizadas em pesquisas e tratamentos de
embriões nas fases de mórula e blastocisto.

Sobre as células tronco embrionárias:
As células tronco, que podem ser classificadas como sendo embrionárias ou adultas, são as células indiferenciadas responsáveis por gerar os outros tipos celulares do organismo e também mais células tronco. Ainda é possível classificar os tipos de células tronco de acordo com o potencial que têm de gerar números de tipos celulares diferentes:
Células tronco totipotentes (tutti, tudo): são capazes de gerar todos os tipos celulares do organismo e inclusive os anexos embrionários. São totipotentes o próprio zigoto e as células tronco embrionárias (blastômeros) do embrião até o estágio de mórula de oito células.
Células tronco pluripotentes: são aquelas capazes de gerar praticamente todos os tipos celulares do organismo, mas não são capazes de gerar os anexos embrionários. Pode-se encontrar esse tipo de célula até a fase de blástula/blastocisto.
Os outros tipos, as células tronco multipotentes por exemplo, são células tronco adultas e são capazes de gerar um número menor de tipos celulares distintos, restringindo-se àqueles presentes nos tecidos onde se encontram.
3.2. Gastrulação: Blástula > Gástrula.
Nesse processo ocorre aumento
do número e do volume das células e é nessa fase que se inicia a diferenciação
celular. No caso do anfioxo, a gastrulação ocorre por invaginação ou embolia: parte
da blástula é dobrada no sentido do interior da blastocela. Ao término da
gastrulação a blastocela desaparece e passa a existir em seu lugar, outra
cavidade, o arquêntero ou gastrocela,
que representa o início da formação do aparelho digestório.
A imagem abaixo mostra uma fatia de embrião, iniciando na fase de blástula até a fase de gástrula. Imagine algo similar a uma fatia de pimentão.
A imagem abaixo mostra uma fatia de embrião, iniciando na fase de blástula até a fase de gástrula. Imagine algo similar a uma fatia de pimentão.



Em relação ao blastóporo os animais podem ser:
Deuterostômios:
o blastóporo origina o ânus, a boca surge posteriormente. Filos dos cordados (o nosso) e dos equinodermas.
Protostômios:
o blastóporo origina a boca, o ânus surge depois. Restante dos filos de
animais, exceto os poríferos.
Os Folhetos germinativos:
São os tecidos embrionários, cuja origem inicia na fase de gástrula, que
posteriormente darão origem aos tecidos adultos. Esses tecidos são três: o ectoderma, o endoderma e
o mesoderma, sendo que o endoderma e o mesoderma originam-se a partir de um tecido embrionário precursor, chamado mesentoderma ou mesendoderma. Os tecidos adultos são o nervoso, os musculares, os epiteliais e os
conjuntivos.
Ectoderma: tecido
localizado mais externamente. Origina o sistema nervoso, a epiderme e os anexos
da pele (pelos, unhas e glândulas), os revestimentos nasal, bucal e anal, e o
esmalte dos dentes (porção mais externa dos dentes).
Endoderma: tecido localizado mais internamente. Epitélio
de revestimento interno do aparelho digestório (exceto o das cavidades oral e
anal, que são originados pelo ectoderma). Origina também o sistema respiratório,
o fígado e o pâncreas.
Mesoderma: tecido localizado entre a ectoderme e a endoderme. Origina: músculos, derme, cartilagens, ossos e outros tecidos
conjuntivos, o sangue e as estruturas do sistema circulatório, e os órgãos dos
sistemas genital e urinário.
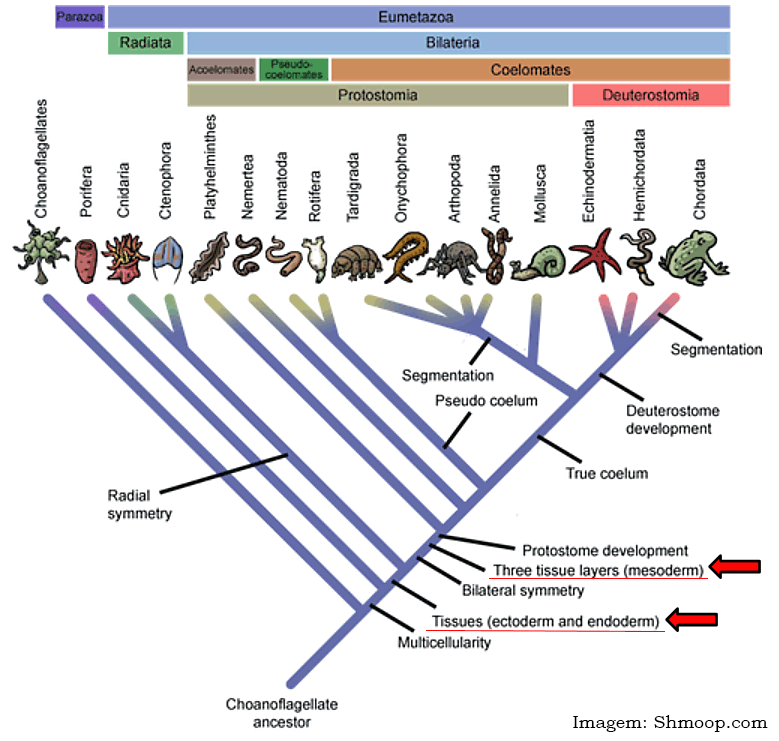
Nem todos os filos de animais possuem os três folhetos germinativos. Por exemplo, as esponjas, Filo Porifera, não possuem folhetos germinativos, pois não possuem tecidos verdadeiros (Parazoa). Na medida em que os animais foram se tornando mais complexos, graças ao processo evolutivo, surgiram os animais com tecidos verdadeiros (Eumetazoa). Inicialmente desenvolveram-se os que têm apenas dois folhetos, a endoderme e a ectoderme, como os do Filo Cnidaria, classificados nesse âmbito como sendo diblásticos. Posteriormente surgiram os triblásticos, esses têm os três folhetos germinativos. Veja o cladograma abaixo.
3.3. Organogênese: Gástrula > Nêurula.
Na fase de nêurula ocorre a formação inicial dos sistemas de órgãos corporais a partir da diferenciação
das células dos folhetos germinativos. Em destaque, são originados nessa etapa
o tubo nervoso dorsal (ectoderma), a notocorda, os somitos e o celoma (mesoderma):
Tubo nervoso dorsal (ectoderma): é o sistema nervoso do animal.
Somitos (mesoderma): estruturas que se repetem ao longo do corpo e que originam, dentre
outras coisas, os músculos e os ossos (nos cordados que possuem ossos).
Celoma:
cavidade corporal completamente revestida por mesoderma onde se localizam
alguns órgãos corporais.
Notocorda (mesoderma): estrutura de sustentação. Também auxilia na formação
do tubo nervoso dorsal. Nos cordados superiores, como os Craniata, a notocorda
é substituída pela coluna vertebral (substituída, ela não “vira” e nem
se transforma na coluna vertebral).
Imagine que na figura abaixo você está vendo uma fatia de anfioxo, a cabeça do animal estaria fora da tela e a cauda dentro.


Imagem mostrando a formação do tubo nervoso dorsal (roxo) em um embrião de animal.
Nem todos os animais triblásticos possuem celoma. No que diz respeito ao celoma os triblásticos podem ser classificados como sendo:
- Acelomados: não possuem celoma, o mesoderma não apresenta cavidade. Exemplo: Filo Platyhelminthes.
- Pseudocelomados: possuem um pseudoceloma, a cavidade existe, mas não é completamente revestida pelo mesoderma. Exemplo: Filo Nematoda.
- Celomados: têm celoma verdadeiro, a cavidade completamente revestida por mesoderma. Filos restantes de animais triblásticos.

4. Os anexos embrionários presentes nos amniotos:
Os anexos embrionários são estruturas
originadas a partir dos folhetos germinativos que auxiliam no desenvolvimento
do embrião, mas não farão parte do corpo do animal. Nem todos os animais apresentam todos os anexos apenas os amniontes, que são os répteis, aves e mamíferos possuem todos os quatro. Aparentemente, os anfioxos têm apenas o saco vitelínico. De qualquer forma, os anexos embrionários são:
Cório: membrana
que recobre todo o embrião e os outros anexos. Responsável pelas trocas gasosas
entre o embrião e o ambiente externo.
Âmnio: membrana
que reveste o embrião e cujo interior é preenchido por liquido (amniótico). Protege
contra a dessecação e os choques mecânicos.
Vesícula vitelínica: bolsa que armazena o vitelo. Participa da nutrição do embrião.
Alantoide: anexo
associado ao intestino do embrião, cuja função principal é a de armazenar os
excretas nitrogenados. Além disso, por ter uma porção ligada ao cório, também
participa das trocas gasosas.

Anexos embrionários de um mamífero (esquerda) e os de uma ave ou réptil (direita).
Veja que o cordão umbilical dos mamíferos (esquerda) é gerada por partes do cório, alantoide e saco vitelínico. Esses dois últimos anexos são vestigiais nos mamíferos placentários, visto que são vivíparos e têm placenta. Os ovos das aves, dos répteis e dos mamíferos monotremados estão dentre os que possuem os anexos embrionários mais bem desenvolvidos.
OBS - Placenta:
é um órgão formado por tecidos do embrião (cório) e da mãe (endométrio), cujas
funções principais são:
I. Permitir a troca de substâncias entre mãe e filho, sem haver mistura de sangue.
- Excretas e CO2: Filho à Mãe.
- Nutrientes e O2: Mãe à Filho.
II. Produção de hormônios:
- hCG (gonadotrofina coriônica humana): Mantém a integridade do corpo amarelo, de forma que ele continue a produzir estrógeno e progesterona. Esses dois hormônios impedem a ovulação e mantém a integridade do endométrio.
- Posteriormente, após a degeneração do corpo amarelo, a placenta passa a produzir estrógeno e progesterona.

REFERÊNCIAS:
Amabis & Martho. Biologia
das Células. 3ª ed. Moderna. 2010.
Bertrand, S. et al. FGFRL1 is a neglected putative actor of the FGF
signalling pathway present in all major metazoan phyla. BMC Evolutionary
Biology. v.9(226). 2009.
Campbell et al. Biologia. 8ª ed.
Artmed. 2010.
Catani, A. et al. Ser
protagonista Biologia – Vol. 1. Edições SM. 2009.
Gilbert & Barresi. Developmental Biology. 11th ed. Sinauer Associates. 2016.
Gilbert & Barresi. Developmental Biology. 11th ed. Sinauer Associates. 2016.
Holland, LZ. & Onai, T. Early development of cephalochordates
(amphioxus). Wires Dev Biol. v. 1. 2012.
Strelchenko, N. & Verlinsky, Y. Embryonic stem cells from morula.
Methods Enzymol. v. 418. 2006. pp: 93-108.
Na internet:
http://www.ucmp.berkeley.edu/vertebrates/tetrapods/amniota.html
http://stemcells.nih.gov
https://ib.bioninja.com.au/standard-level/topic-1-cell-biology/11-introduction-to-cells/stem-cells-2.html
https://ib.bioninja.com.au/standard-level/topic-1-cell-biology/11-introduction-to-cells/stem-cells-2.html
https://www.cell.com/fulltext/S0092-8674(01)00307-5
https://www.nature.com/articles/s41467-020-17575-w
https://www.frontiersin.org/articles/10.3389/fcell.2021.668006/full
Wikipedia. Entradas diversas sobre
o tema.










{kind=link}